FOCUS
Fisiologia di apprendimento e memoria
Mariano Pedetti – SerT MVT, AUSL2 dell‘Umbria, Marsciano (PG)

L‘apprendimento è la capacità del cervello di imparare dall‘esperienza, cioè di registrare nella memoria, o meglio nelle memorie, tutto ciò che può essere utile in futuro per le successive interazioni dell‘individuo con l‘ambiente. Nonostante esistano periodi critici per particolari forme di apprendimento (per esempio, la finestra linguistica) questo dura per tutta la vita, grazie alla capacità del cervello di riaggiornare continuamente i suoi repertori mnesici (neuroplasticità).
Tipologia delle memorie
<div “margin:8px;”=””>

La memoria non è un fenomeno unitario; al contrario, le memorie sono molteplici e codificate, e conservate in reti neurali ampiamente distribuite in varie regioni del cervello (Fig. 1).
La distinzione fondamentale della memoria umana è fra quella dichiarativa, o esplicita, tipica degli esseri umani in quanto necessita del linguaggio simbolico, e quella non dichiarativa , o implicita, presente a vari livelli di complessità in tutte le forme viventi (Fig. 2).(1)
Il sistema mnesico del lobo temporale (formato dalla corteccia paraippocampale e dall‘ippocampo) si occupa della codifica delle memorie dichiarative o esplicite (così definite perché possono essere recuperate consciamente dalla memoria di lavoro), suddivise in episodiche (eventi di cui abbiamo fatto esperienza in prima persona) e fattuali (o semantiche). L‘ippocampo genera anche le memorie spaziali o di contesto, probabilmente all‘interno di una più generale funzione di formazione delle memorie relazionali. I ricordi dichiarativi rimangono immagazzinati nell‘ippocampo solo a breve-medio termine. In un arco temporale variabile fra uno-due anni, le memorie sono trasferite e distribuite in varie regioni corticali (memoria a lungo termine), quelle dove sono state codificate le rappresentazioni. I ricordi visivi nella corteccia visiva, gli uditivi in quella uditiva ecc.

La memoria di lavoro, o memoria operativa o memory working, può essere definita un database temporaneo (memoria a brevissimo termine) per la conservazione delle informazioni per il tempo necessario a completare un‘azione. Lo span è la capacità della memoria di lavoro, cioè il numero di elementi che può utilizzare a un dato istante, in genere sette. La memoria di lavoro, distribuita in subregioni della CPF, soprattutto nella CPF dorsolaterale, è il trait d‘union fra la dimensione percettiva e quella cognitiva cosciente del cervello. Subsistemi separati di memory working inviano, dalle aree corticali sensoriali (visiva, uditiva ecc.), informazioni alla CPF dorsolaterale, che le integra in modalità multimodale.(2) La dopamina modula le funzioni della memoria di lavoro influenzando l‘attenzione selettiva: la stimolazione dei recettori corticali del sottotipo D1-like (D1-D5), postsinaptici, ad alta affinità di legame, si traduce in una facilitazione dei pattern di scarica dei neuroni che devono lavorare, riconosciuti in base al loro stato elettrico, mentre quelli non utili al momento vengono inibiti dalla stimolazione dei recettori presinaptici a bassa affinità, del sottotipo D2-like (D2-D3-D4).(3) In sintesi, la dopamina “dosa” il numero d‘impulsi generati nelle aree prefrontali della memory working e provocati dalle informazioni provenienti dalle regioni sensoriali. Quando il compito è stato svolto, output inibitori dai neuroni dei sistemi motori, sia corticali sia subcorticali, inibiscono le cellule dopaminergiche della VTA. Avete mai studiato in un ambiente rumoroso, per esempio in treno? Certamente sì. Ebbene, ci siete riusciti grazie a questa capacità del cervello!
È poi interessante notare che la memoria di lavoro, in quanto collo di bottiglia da cui passa tutta la comunicazione simbolica interpersonale, è anche il ricevitore corticale della terapia psicologica verbale. I sistemi mnesici impliciti sono svincolati dal lobo temporale; infatti, funzionano anche in caso di lesione di questo. Tuttavia, forniscono anch‘essi informazioni alla memoria di lavoro, per esempio circa gli stati del corpo tramite l‘insula, o le reazioni apprese di paura codificate dall‘amigdala, e ricevono connessioni dalla CPF.(4) Il priming è un tipo particolare di apprendimento implicito, che ha preso il nome da un test usato negli amnesici ippocampali. Se a uno di questi soggetti si fa vedere una lista di tre parole, per esempio pane, finestra e crotalo, e il giorno successivo si chiede loro di ripetere la lista, non solo non ricordano le parole, ma nemmeno di aver visto la lista (amnesia anterograda). Tuttavia, se si sillaba cro-, loro associano la parola crotalo e non crosta o crollo. (3)

Le tipologie di memorie implicite per noi più interessanti sono quelle formate tramite apprendimento incentivo. Alla base dell‘apprendimento incentivo vi sono il paradigma dell‘apprendimento classico, o pavloviano, e quello del comportamento operante, o strumentale, o skinneriano.(2) Nel primo, un animale può essere addestrato (condizionato) a esprimere una risposta automatica al presentarsi di uno stimolo neutro, come una luce o un suono, dopo un certo numero di associazioni con uno stimolo incondizionato appetibile o aversivo (per esempio, cibo o scossa elettrica) che segue di un breve intervallo temporale la presentazione dello stimolo neutro. Nel caso del cane di Pavlov, lo stimolo incondizionato era un cibo appetibile, la risposta incondizionata la salivazione; dopo un certo numero di presentazioni di un suono in stretta associazione temporale con il cibo, il cane salivava alla presentazione del suono senza cibo (stimolo condizionato). Non sono create nuove risposte, l‘apprendimento consiste nell‘associare una risposta innata a circostanze nuove. Dopo un certo numero di presentazioni dello stimolo condizionato (suono) senza stimolo incondizionato (cibo), la risposta condizionata si indebolisce, fino ad estinguersi del tutto (estinzione del condizionamento). L‘estinzione, però, non cancella l‘associazione suono-cibo dalla memoria, l‘estinzione è in realtà un nuovo apprendimento che inibisce il precedente. Infatti, a distanza di tempo dall‘estinzione al ripresentarsi dello stimolo condizionato la risposta condizionata può ricomparire (fenomeno chiamato recupero spontaneo), mentre la generalizzazione è l‘attivazione della risposta in seguito all‘esposizione a stimoli simili a quello condizionato.
Nel paradigma del comportamento operante o strumentale, o skinneriano, un animale è addestrato a compiere un lavoro o ad attuare un comportamento, anche complesso, nuovo, non compreso nel suo repertorio innato, alla presenza di stimoli discriminativi associati a una ricompensa (rinforzo positivo) o a una punizione (rinforzo negativo). Con l‘instaurarsi del condizionamento gli stimoli discriminativi diventano stimoli condizionati (o rinforzi condizionati), capaci di innescare il comportamento in assenza dei rinforzi incondizionati (ricompensa o punizione). Anche nel paradigma operante si verificano i fenomeni dell‘estinzione del condizionamento, del recupero spontaneo e della generalizzazione.(2)
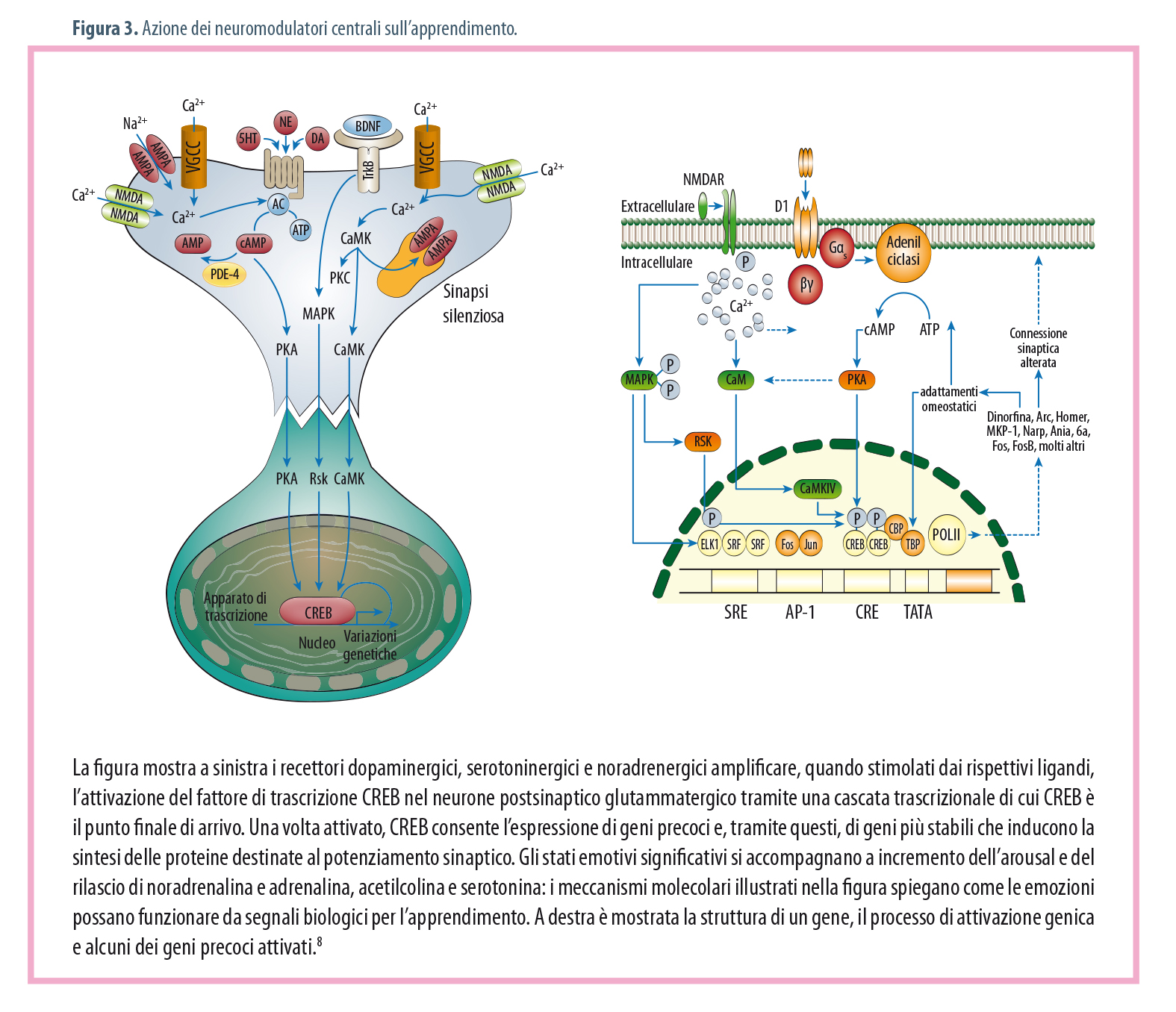
Il PIT, Pavlovian Instrumental Transfer, è un fenomeno molto importante per l‘apprendimento implicito e consiste in un trasferimento di segnale, nel senso che facendo precedere o seguire un apprendimento pavloviano a uno operante, dopo un certo numero di associazioni, lo stimolo condizionato (CS) pavloviano è in grado di avviare e intensificare il comportamento strumentale.(5) Gran parte dell‘apprendimento è di tipo incentivato, anche quello più elevato, come l‘interiorizzazione delle norme etiche, e implicito. La base neurale è il sistema mnesico centrato sull‘amigdala e altre strutture limbiche funzionalmente correlate fra loro e deputate a elaborazioni emotivo-istintuali. L‘apprendimento incentivo consente l‘associazione e la memorizzazione di sensazioni piacevoli (ricercate attivamente) e spiacevoli (evitate attivamente) con eventi, fatti, luoghi, contesti ambientali. Funziona tanto meglio quanto più le sensazioni sono vivide e intense, emotivamente salienti e importanti per la sopravvivenza (Figg. 3 e 4).
Determina la codifica di schemi comportamentali che il cervello ritiene dover ripetere con massima efficienza, in quanto pro-fitness (per fitness si intendono le attività a maggior vantaggio evolutivo, cioè quelle con la massima probabilità di conservare e trasmettere il patrimonio genetico specie-specifico).(6)
Il cervello matura le diverse funzioni mnesiche in tempi successivi: lo sviluppo ontogenetico ripercorre le tappe di quello filogenetico, per cui i sistemi mnesici impliciti, subcorticali, maturano prima di quelli espliciti, corticali.
Tutti impariamo un sacco di cose inconsapevolmente, prima di aver imparato a leggere e a scrivere e di essere capaci di formare memorie dichiarative.(1) Un esempio è la strutturazione del repertorio affettivo-emotivo in relazione alle cure parentali ricevute nei primi due-tre anni di vita, di cui non abbiamo ricordo. L‘apprendimento implicito continua per tutta la vita. Sappiamo qualcosa sui meccanismi biologici alla base di tutto ciò? Come al solito molto, e ancora troppo poco.
Plasticità hebbiana, PLT e DLT
La maggior parte dei neuroscienziati ritiene che modificazioni nella connettività sinaptica siano alla base dell‘apprendimento, e che la memoria sia il consolidamento e la conservazione di questi cambiamenti nel tempo.
Quali sono questi cambiamenti?

Un primo passo fu fatto da Donald Hebb, con la sua teoria sinaptica della memoria o della connessione tramite scarica: se due neuroni sono attivi nello stesso istante, e uno è in posizione presinaptica rispetto all‘altro, allora la connessione fra loro ne sarà potenziata. In altri termini, cellule che scaricano in simultanea si connettono reciprocamente. Immaginiamo di avere tre neuroni A, B, C: A è in posizione postsinaptica rispetto a B e C, e la connessione fra A e C è forte, mentre quella fra A e B è debole. Quando C scarica, anche in A si forma un potenziale d‘azione, mentre è meno probabile che si verifichi quando scarica B. B e C sono impegnati nell‘elaborazione di stimoli diversi: a un dato momento scaricano insieme, e A si attiva. Secondo il principio di Hebb, la connessione A-B è stata rafforzata dalla scarica sincrona di A-B-C. L‘intensità del rafforzamento correla direttamente con la frequenza delle scariche sincrone, fino a che C sarà in grado di attivare A senza l‘intervento di B. (2)
Quasi 25 anni dopo, l‘ipotesi di Hebb fu dimostrata sperimentalmente in uno studio sull‘ippocampo del ratto.<div “margin:5px;”=””>

I ricercatori impiantarono un elettrodo di registrazione nell‘ippocampo e uno nella via nervosa che raggiungeva l‘ippocampo, misurarono l‘attività elettrica dopo uno stimolo unico, poi dopo una breve scarica ad alta frequenza, e continuarono a farlo per ore. La scoperta, che prese il nome di potenziamento a lungo termine (PLT), fu che dopo gli stimoli di potenziamento la risposta sinaptica aumentava e si manteneva nel tempo.(2)
Da allora sono stati pubblicati migliaia di studi sul PLT e il suo opposto, la DLT (depressione a lungo termine). I risultati, ai nostri fini, possono essere schematicamente sintetizzati così: il PLT è specifico per le sinapsi coinvolte dal potenziamento e non per il neurone nel suo complesso (cioè si forma solo nelle sinapsi stimolate e non in tutte le sinapsi del neurone) ed è associativo, si verifica in tutte le sinapsi attive al momento del potenziamento.
La sinapsi glutammatergica
La risposta alla domanda riguardo a come il cervello realizza la plasticità hebbiana ha impegnato i neuroscienziati per molti anni, e ha rappresentato la linea di partenza per gli studi su apprendimento e memoria. A metà degli anni Ottanta furono fatte due scoperte fondamentali, che hanno rappresentato uno starter per gli studi successivi:
• il blocco del recettore NMDA del glutammato impedisce il PLT senza interferire con la normale trasmissione sinaptica;
• il blocco dell‘ingresso di calcio nella cellula postsinaptica durante il potenziale di azione impedisce il PLT.
Gli studi successivi hanno definito il funzionamento della sinapsi glutammatergica, illustrato dalla Figura 5.

Il glutammato ha parecchi recettori (Box 1), tuttavia, per la nostra discussione, sono essenziali i recettori AMPA, responsabili della trasmissione sinaptica ordinaria, e i recettori NMDA, coinvolti nella plasticità sinaptica. La Figura 5A rappresenta nella parte superiore una connessione forte e in quella inferiore una connessione debole allo stesso neurone postsinaptico. Il glutammato rilasciato nello spazio presinaptico raggiunge sia i recettori AMPA sia quelli NMDA: inizialmente sul recettore NMDA non ha effetto, in quanto questo è bloccato dal Mg, mentre il legame con quello AMPA induce un potenziale di azione nella cellula postsinaptica (connessione forte). Ora, se le due connessioni presinaptiche scaricano insieme, il potenziale d‘azione indotto da quella forte rimuove il Mg dal recettore NMDA, a cui segue l‘ingresso di calcio attraverso il canale NMDA, inducendo un PLT precoce. A ciò si associa l‘avvio di modificazioni intracellulari che contribuiscono al potenziamento sinaptico, utilizzando le proteine già presenti nel citosol (memoria a breve o brevissimo termine). I recettori NMDA funzionano da rilevatori di coincidenza, permettendo al neurone postsinaptico di registrare esattamente quali input presinaptici fossero attivi durante il suo potenziale d‘azione.
Ora, bisogna capire in che modo le modificazioni sinaptiche indotte dall‘attività NMDA si consolidino in modo da generare PLT tardivi e, tramite fenomeni stabili di plasticità neurale, le memorie a medio e lungo termine. Il primum movens è l‘ingresso di ioni calcio nella cellula, dove si comportano da secondo messaggero, controllando e indirizzando reazioni chimiche che potenziano le connessioni sinaptiche sia a breve sia a lungo termine. L‘ingresso di calcio attiva diverse chinasi a cascata: la proteinchinasi A cAMP-dipendente (PKA), la proteinchinasi calcio/calmodulina (CaMK), la proteinchinasi attivata da mitogeno (MAPK). Tutte le chinasi suddette convergono, a loro volta, nell‘attivazione del fattore di trascrizione genica CREB (elemento di risposta al cAMP o regione di legame proteico), che agisce sui segmenti promoter dei geni, attivandone l‘espressione e, quindi, la sintesi proteica. Le proteine sintetizzate sono trasportate lungo la cellula, ma solo le sinapsi marcate durante il PLT mediante messaggeri retrogradi rilasciati dal neurone postsinaptico potranno utilizzarle, e quindi potenziarsi. Le proteine servono per costruire nuovi recettori, nuovi canali ionici, ma non solo: si ha anche la formazione di nuove sinapsi.
Alcune delle proteine rilasciate antidromicamente sono neurotrofine, che sono catturate dai terminali sinaptici attivi in quel momento e sollecitano la cellula presinaptica a produrre nuove connessioni sinaptiche con i neuroni postsinaptici. Il DLT è il fenomeno opposto del PLT: si verifica quando una via presinaptica scarica ripetutamente non in coincidenza con il neurone postsinaptico.
<div “margin:8px;”=””>

In questo caso, lo scarso afflusso di calcio induce l‘attivazione di proteinfosfatasi che defosforilano i recettori AMPA e, in questo modo, depotenziano il collegamento sinaptico. Il calcio è il regolatore bidirezionale della plasticità.(2)
Le sinapsi si potenziano e depotenziano, nascono e muoiono in continuazione, e questa caratteristica, che come abbiamo già detto è definita nel suo complesso neuroplasticità, è la base della nostra straordinaria capacità di adattamento alle mutevoli esigenze dell‘ambiente.
Tuttavia, a volte le memorie sono particolarmente durature, alcune muoiono con noi. Una possibile risposta potrebbe essere la recente scoperta, in una lumaca di mare, di CPEB (proteina che si lega al fattore citoplasmatico di poliadenilazione). Ha una struttura simile ai prioni (nella forma autosomica dominante si autoperpetua), è attivata dalla serotonina e sostiene una sintesi proteica perpetua nelle sinapsi, mantenendo il rimodellamento sinaptico alla base dell‘apprendimento a lungo termine.(1)
Dato che i meccanismi fondamentali per la vita sono mantenuti dall‘evoluzione in maniera simile nei vari organismi viventi, indipendentemente dalla loro complessità, questa scoperta ha aperto un nuovo filone della biologia, che potrebbe decifrare il mistero molecolare delle memorie indelebili anche nei mammiferi, uomo compreso.
È infine doveroso ricordare che esistono diversi tipi di PLT e DLT, oltre a quello sinteticamente descritto (il più comune e conosciuto), e che PLT e DLT non sono esclusiva dei neuroni ippocampali, ma sono stati individuati in tutte le regioni cerebrali. In conclusione, i ricordi sono la conservazione degli apprendimenti; non sono fotogrammi statici, ma al contrario rappresentazioni dinamiche (il passato è presente ricordato), ogni volta diverse dalle precedenti.
Sono codificati sotto forma di pattern di scarica elettrica in circuiti distribuiti e interconnessi, sottoposti a continui cambiamenti sotto le spinte di nuove esperienze, dei contesti in cui si verificano e dell‘arousal emotivo che le accompagna. Ogni nuovo apprendimento espande il repertorio mentale dell‘individuo e quindi, impercettibilmente ma continuamente, modifica l‘influenza reciproca fra memoria e apprendimento.
Bibliografia
1. E.R. Kandel, in “Alla ricerca della memoria”, Codice, Torino, 2010.
2. J. LeDoux, in ” Il Se sinaptico”, Raffaello Cortina, Milano, 2001.
3. D.J. Linden, in “La mente casuale”, Centro Scientifico Editore, Torino, 2009.
4. L. Clark, et al., “Differential effects of insular and ventromedial prefrontal cortex lesions on risky decision-making”, Brain 2008;131(Pt 5):1311-22.
5. B.J. Everitt, T.W. Robbins, “Neural systems of reinforcement for drug addiction: from actions to habits to compulsion”, Nat Neurosci 2005;8(11):1481-9.
6. R. Montague, in “Perche l‘hai fatto?”, Raffaello Cortina, Milano, 2008.
7. http://www.ferraldo.eu/didattica/M&C1/Col%2005)%20Neuromediatori%20e%20recettori.pdf
8. C.A. McClung, et al., “DeltaFosB: a molecular switch for long-term adaptation in the brain”, Brain Res Mol Brain Res
2004;132(2):146-54.
9. W. Schultz, “Multiple dopamine functions at different time courses”, Annu Rev Neurosci 2007;30:259-88.